STEMdiff™ Cerebral Organoid Maturation Kit
Culture medium kit for extended maturation of human cerebral organoids

Request Pricing
Thank you for your interest in this product. Please provide us with your contact information and your local representative will contact you with a customized quote. Where appropriate, they can also assist you with a(n):
Estimated delivery time for your area
Product sample or exclusive offer
In-lab demonstration
This site is protected by reCAPTCHA and the Ěý˛ą˛Ô»ĺĚýĚý˛ą±č±č±ô˛â.
- Ultra-Low Adherent Plate for Suspension Cultu...
Sterile, flat-bottom plate, with ultra-low adherent surface for suspension cultures, with lid; 6-well format
- Y-27632 (Dihydrochloride)
RHO/ROCK pathway inhibitor; Inhibits ROCK1 and ROCK2
What Our Scientist Says
Human brain development is complex, so to observe it in a dish is a huge scientific breakthrough. We've tried to simplify that process and make it more accessible to you, regardless of how much stem cell experience you have.

Overview
Data Figures
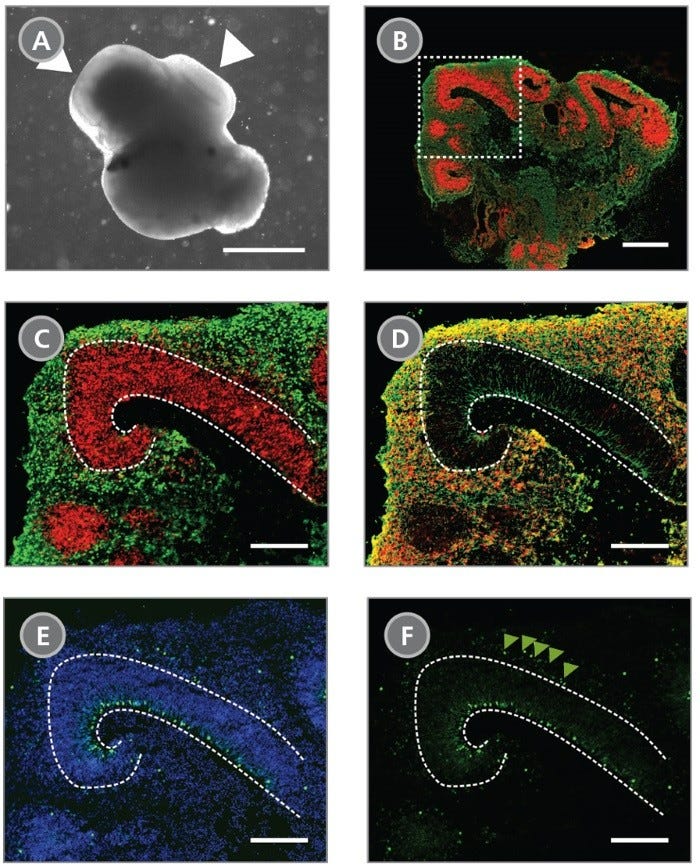
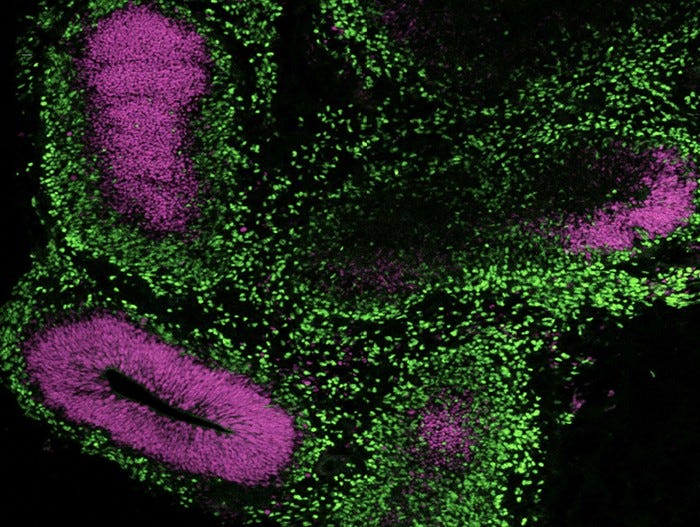
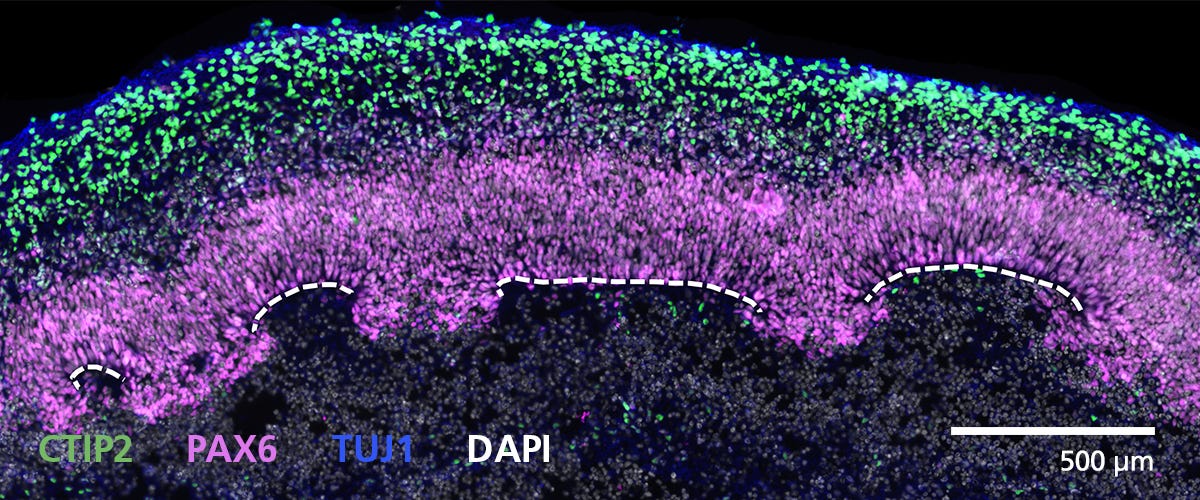
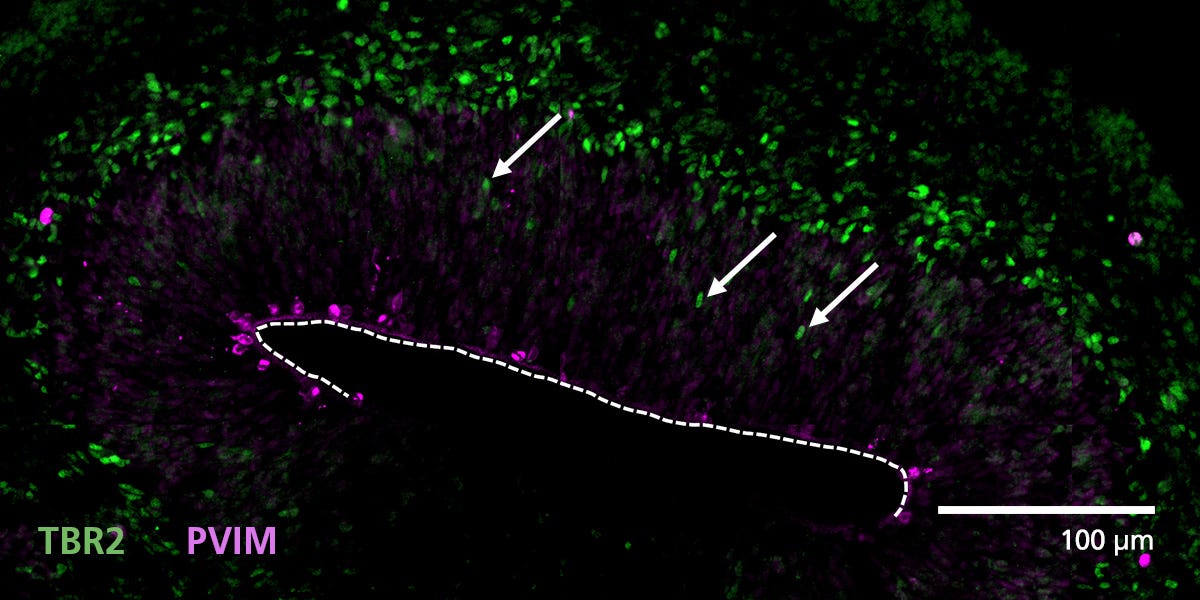
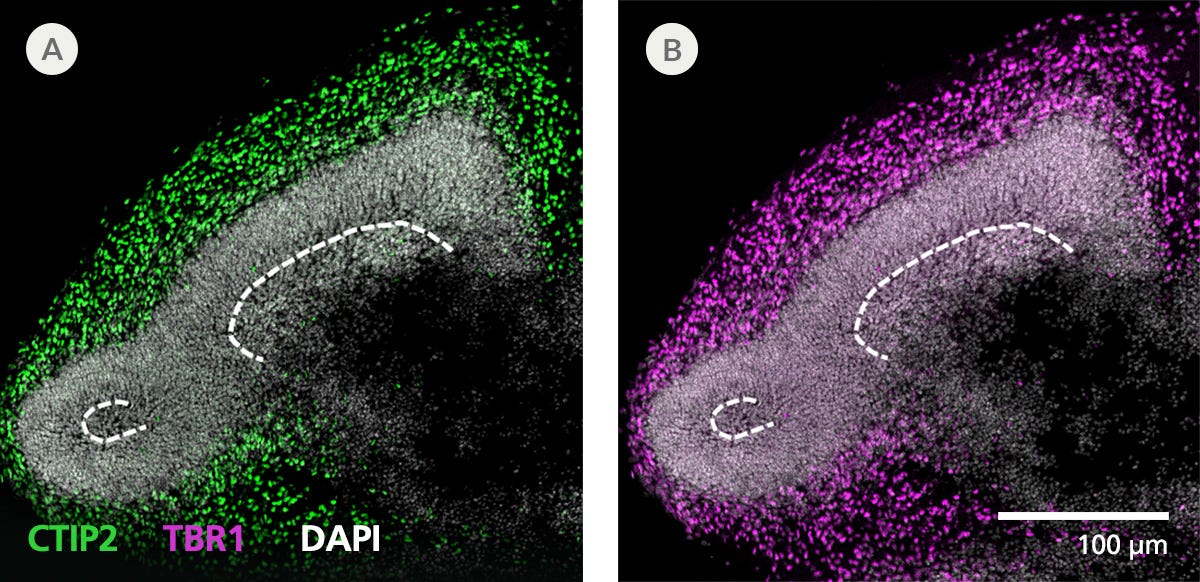
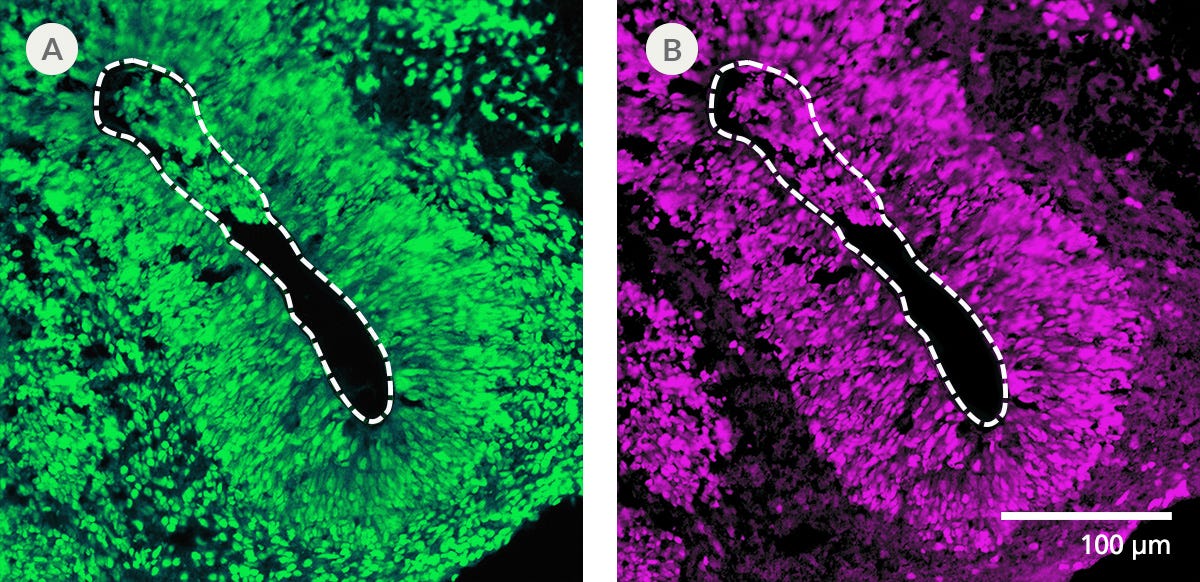
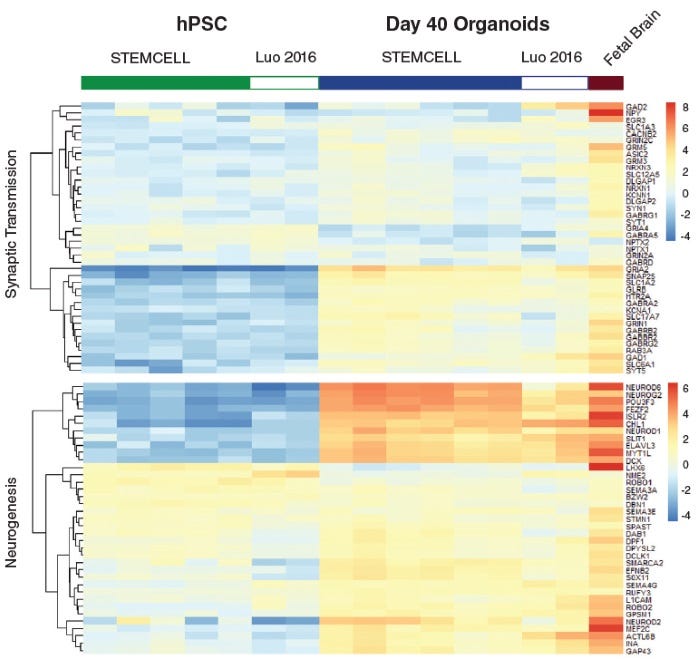
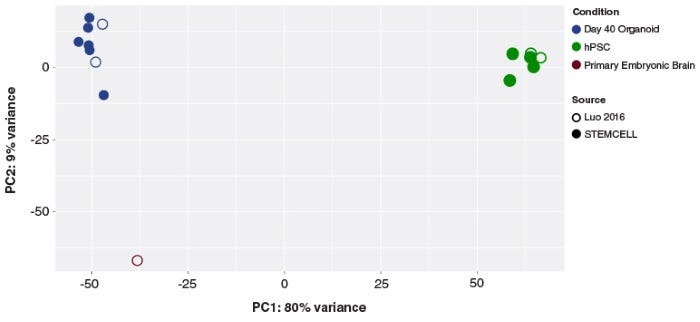

Protocols and Documentation
Find supporting information and directions for use in the Product Information Sheet or explore additional protocols below.
Applications
This product is designed for use in the following research area(s) as part of the highlighted workflow stage(s). Explore these workflows to learn more about the other products we offer to support each research area.
Resources and Publications
Educational Materials (41)

































Publications (12)
Abstract
Abstract
Abstract
Related Products
This product was developed under a license to intellectual property owned by the Institute of Molecular Biotechnology (IMBA) of the Austrian Academy of Sciences. This product is sold for research use only (whether the buyer is an academic or for-profit entity) under a non-transferable, limited-use license. Purchase of this product does not include the right to sell, use or otherwise transfer this product for commercial purposes (i.e., any activity undertaken for consideration, such as use of this product for manufacturing, or resale of this product or any materials made using this product, or use of this product or any materials made using this product to provide services or, in collaboration with, a for-profit entity, for purposes other than research applications (i.e., drug development activities). Purchasers wishing to use the product for commercial purposes should contact IMBA at technologvtransfer@imba.oeaw.ac.at.
PRODUCTS ARE FOR RESEARCH USE ONLY AND NOT INTENDED FOR HUMAN OR ANIMAL DIAGNOSTIC OR THERAPEUTIC USES UNLESS OTHERWISE STATED. FOR ADDITIONAL INFORMATION ON QUALITY AT şŁ˝ÇĆĆ˝â°ć, REFER TO WWW.şŁ˝ÇĆĆ˝â°ć.COM/COMPLIANCE.
CA WARNING: This product can expose you to Progesterone which is known to the State of California to cause cancer. For more information go to
